Louvain Drug Research Institute > Cellular and Molecular Pharmacology
|
|
Université
catholique de Louvain Louvain Drug Research Institute > Cellular and Molecular Pharmacology |
|
|
Cellular toxicity of antibiotics |
|
Quick
links
|
Understanding the
cellular and molecular mechanisms of antibiotic toxicity is critical for
improving our therapeutic arsenal and select drugs with the best benefit
to risk ratio. We focus our attention on the cellular lipidosis and apoptosis induced by antibiotics, and try to understand the corresponding underlying mechanisms, using a combination of biochemical, morphological and biophysical techniques. These research programs are closely linked to those exploring drug-membrane interactions and intracellular chemotherapy (pharmacokinetics) |
|
|
|
|
|
|
|
|
|
|
|
Team
- Senior investigators: M.P. Mingeot-Leclercq, F. Van Bambeke, P.M. Tulkens
- Post-doctoral fellows: D. Das (2010-2012)
- Doctoral fellows: S. Denamur (2007- ), J. Lorent (2007- )
- Former investigators: M.B. Carlier (1984-1988), L. Giurgea (1984-1987), S. Ibrahim (1984-1990), J. Piret (1989-1994), D. Beauchamp (1984-1986), P. Lambricht (1986-1991), R. Wagner (1987-1990), B.K. Kishore (1989-1993), L. Fuming (1990), J.P. Montenez (1991-1995), M. El Mouedden (1994-2000), C. Gerbaux (1994-1999), D. Tyteca (1997-2002), H. Servais (2002-2006), E. Charvalos (2003), H. Aly (2007-2008)
Collaborations
- Prof. P.J. Courtoy & D. Tyteca (Endocytosis and epithelial differentiation, de Duve Institute & Université catholique de Louvain, Brussels).
- Prof. R. Brasseur (Centre de biophysique moléculaire numérique, Université de Liège, Gembloux Agro Biotech, Gembloux)
Main current research programs
The accumulation of antibiotics in lysosomes (by endocytosis or proton trapping) may cause metabolic alterations that can lead to cell toxicity. Three classes of antibiotics have been especially studied in this context.
1. Aminoglycosides
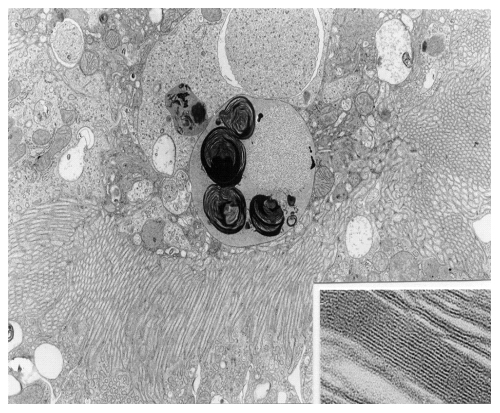
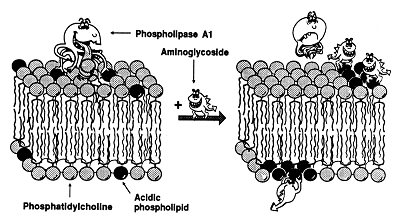
Aminoglycoside antibiotics were amongst the first antibiotics for which a lysosomal accumulation was demonstrated both in cultured cells and in renal proximal tubular cells in vivo. These antibiotics cause a lysosomal phospholipidosis (see Figure 1A) which we consider to be a hallmark of their renal toxicity. Phospholipidosis is the consequence of the binding of these cationic antibiotics to negatively-charged phospholipids in membrane, which is favored by the acidic pH of lysosomes. In vitro studies have shown that negative charges are essential for the activity of lysosomal phospholipases. The binding of aminglycosides to phospholipid bilayers in lysosomes may, therefore, results in a decreased activity of the phospholipases, which explains the accumulation of undegraded phospholipids (see model in Figure 1B).
In addition, aminoglycosides cause membrane permeabilization as well as membrane bilayer aggregation, and impair membrane fusion.
|
These studies are being extended to the evaluation of novel molecules with reduced binding to negatively-charged phospholipids and/or reduced accumulation by renal proximal tubular cells.
2. Macrolides and lipoglycopeptides
Macrolides also accumulate in lysosomes (by proton trapping), whereas the novel lipoglycopeptides (telavancin, oritavancin) enter lysosomes by adsorptive endocytosis.
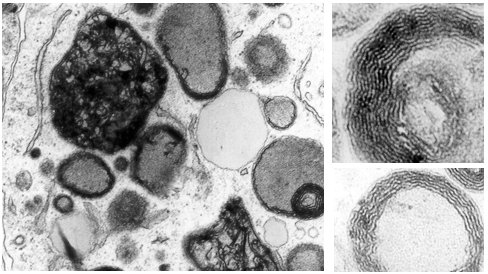
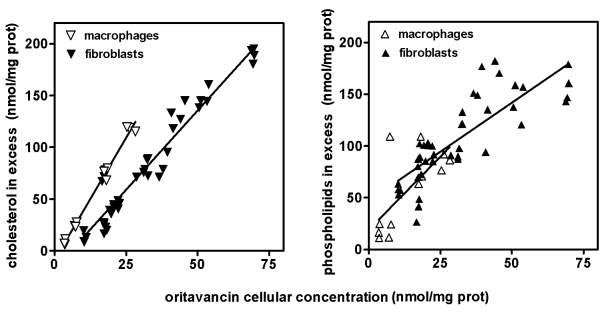
By interacting with phospholipids, macrolides and lipoglycopeptides (especially those with a large cellular accumulation) induce a lysosomal phospholidosis (see Figure 2A). However, and in contrast with what has been described for aminoglycosides (see above), they also cause an accumulation of cholesterol and, probably, other lipids. This is particularly conspicuous in cells exposed to lipoglycopeptides (see Figure 2B). Similar alterations are seen in animals treated with large doses of these antibiotics.
|
These studies are now extended to new molecules as a basis for their pre-clinical assessment for early toxicity related to perturbations of lipid metabolism.
Selected References on drug-induced lipidosis (by reverse chronological order; for a full reference list, see our publication list)
Aminoglycosides
- Servais H, Mingeot-Leclercq MP, and Tulkens PM 2004. Antibiotic-induced nephrotoxicity In: Toxicology of the Kidney (Target Organ Toxicology Series), chap. 16, pp 635-685, J.B. Tarloff & L.H. Lash, eds., CRC Press, Boca Raton, Fla., ISBN 0-415-24864-7 (PDF)
- Mingeot-Leclercq MP, Tulkens PM. 1999. Aminoglycosides: nephrotoxicity. Antimicrob. Agents Chemother. 43:1003-1012. (PDF)
- Van Bambeke, F., P.M. Tulkens, R. Brasseur & M.-P. Mingeot-Leclercq. 1995. Aminoglycoside antibiotics induce aggregation but not fusion of negatively-charged liposomes. Eur. J. Pharmacol. 289:321-333.
- Kotretsou, S., M.-P. Mingeot-Leclercq, V. Constantinou-Kokotou, R. Brasseur, M.P. Georgiadis & P.M. Tulkens. 1995. Synthesis and antimicrobial and toxicological studies of amino acid and peptide derivatives of kanamycin A and netilmicin. J. Med. Chem. 38:4710-4719
- Van Bambeke, F., M.P. Mingeot-Leclercq, A. Schanck, R. Brasseur & P.M. Tulkens. 1993. Alterations in membrane permeability induced by aminoglycoside antibiotics: studies on liposomes and cultured cells. Eur. J. Pharmacol. 247:155-168
- Mingeot-Leclercq, M.P., A. Van Schepdael, R. Brasseur, R. Busson, H.J. Vanderhaeghe, P.J. Claes & P.M. Tulkens. 1991. New derivatives of Kanamycin B obtained by modifications and substitutions in position 6". II. In vitro and computer-aided toxicological evaluation, with respect to interactions with phosphatidylinositol. J. Med. Chem. 34:1476-1482
- Van Schepdael, A., J. Delcourt, M. Mulier, R. Busson, M.P. Mingeot-Leclercq, P.M. Tulkens & P.J. Claes. 1991. New derivatives of kanamycin B obtained by modifications and substitutions in position 6". I. Synthesis and microbiological evaluation. J. Med. Chem. 34:1468-1475
- Van Schepdael, A., R. Busson, L. Verbist, H.J. Vanderhaeghe, M.P. Mingeot-Leclercq, R. Brasseur & P.M. Tulkens. 1991. Synthesis, antibacterial activity and toxicological evaluation (in vitro and computer-aided) of 1-C-hydroxymethyl kanamycin B and 1-C-hydroxymethyl, 6"chlorokanamycin B. J. Med. Chem. 34:1483-1492
- Mingeot-Leclercq, M.P., J. Piret, P.M. Tulkens & R. Brasseur. 1990. Effect of acidic phospholipids on the activity of lysosomal phospholipases and on their inhibition by aminoglycoside antibiotics. II. Conformational analysis. Biochem. Pharmacol. 40:499-506
- Mingeot-Leclercq, M.P., J. Piret, R. Brasseur & P.M. Tulkens. 1990. Effect of acidic phospholipids on the activity of lysosomal phospholipases and on their inhibition by aminoglycoside antibiotics. I. Biochemical analysis. Biochem. Pharmacol. 40:489-497
- Mingeot-Leclercq, M.P., A. Schanck, M.F. Ronveaux-Dupal, M. Deleers, R. Brasseur, J.M. Ruysschaert & P.M. Tulkens. 1989. Ultrastructural, physico-chemical and conformational study of the interactions of gentamicin and bis(beta-diethylaminoethylether)hexestrol with negatively charged phospholipid bilayers. Biochem. Pharmacol. 38:729-741
- Mingeot-Leclercq, M.P., G. Laurent & P.M. Tulkens. 1988. Biochemical mechanism of aminoglycoside-induced inhibition of phosphatidylcholine hydrolysis by lysosomal phospholipases. Biochem. Pharmacol. 37:591-599
- Tulkens PM. 1986. Experimental studies on nephrotoxicity of aminoglycosides at low doses. Mechanisms and perspectives. Am J Med. 80(6B):105-14.
Macrolides
- Montenez, J.P., F. Van Bambeke, J. Piret, R. Brasseur, P.M. Tulkens & M.P. Mingeot-Leclercq. 1999. Interactions of macrolide antibiotics (erythromycin A, roxithromycin, erythromycylamine and azithromycin) with phospholipids: comparative studies with cultured cells, a-cellular systems, and computer-aided conformational approaches. Toxicol. Appl. Pharm. 156:129-140
- Van Bambeke F, C. Gerbaux, J.M. Michot, M. Bouvier d'Yvoire, J.P. Montenez, P.M. Tulkens. 1998. Lysosomal alterations induced in cultured rat fibroblasts by long-term exposure to low concentrations of azithromycin. J Antimicrob Chemother. 42: 761-76 (PDF)
- Van Bambeke F., J.P. Montenez, J. Piret, P.M. Tulkens, P.J. Courtoy, M.P. Mingeot-Leclercq. 1996. Interaction of the macrolide azithromycin with phospholipids. I. Inhibition of lysosomal phospholipase A1 activity. Eur J Pharmacol. 314: 203-21
Lipoglycopeptides
- Barcia-Macay M., F. Mouaden, M.P. Mingeot-Leclercq, P.M. Tulkens, F. Van Bambeke. 2008. Cellular pharmacokinetics of telavancin, a novel lipoglycopeptide antibiotic, and analysis of lysosomal changes in cultured eukaryotic cells (J774 mouse macrophages; rat embryonic fibroblasts). J. Antimicrob. Chemother. 61:1288-1294. (PDF)
- Van Bambeke F., J. Saffran, M.-P. Mingeot-Leclercq & P.M. Tulkens. 2005. Mixed lipid storage disorder induced in macrophages and fibroblasts by oritavancin (LY333328), a new glycopeptide antibiotic with exceptional cellular accumulation. Antimicrob. Agents Chemother. 49: 1695-1700. (PDF)
Molecular mechanisms of aminoglycoside-induced apoptosis
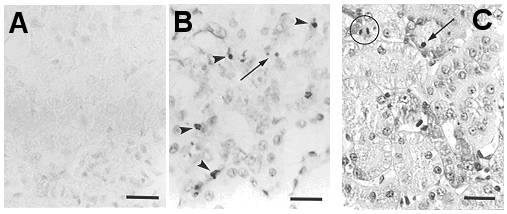
Beside phospholipidosis, aminoglycoside antibiotics also induce apoptosis. Animals treated with low, therapeutically relevant doses of aminoglycosides show both lysosomal phospholipidosis and apoptosis in proximal tubular cells (see Figure 3A). Apoptosis induced by aminoglycosides has been reproduced in vitro with LLC-PK1 and MDCK cells and found to be directly related to the amount of drug accumulated by the cells.
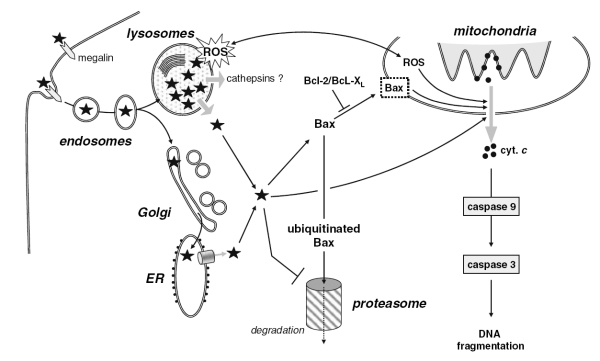
Our current data have shown that gentamicin destabilizes the lysosomal membrane, which could result in the release of the drug and lysosomal constituents such as cathepsins to the cytosol. In this context, the storage of gentamicin in lysosomes would actually appear as a protective mechanism rather than a toxic event, as long as the drug is prevented from moving from there to the cytosol.
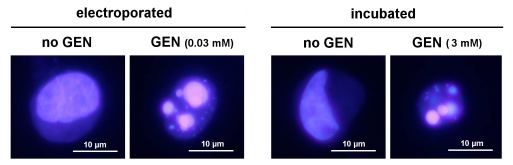
The role of cytosolic gentamicin in triggering apoptosis was further documented by showing that its direct delivery to cells by electroporation induces apoptosis at much lower concentrations than what is required for cells incubated with the drug (see Figure 3B; electroporation can be used for fast screening of potentially less toxic aminoglycosides, as it requires only very low amounts of drugs). In parallel, ongoing reseach examines how gentamicin accumulated in lysosomes could cause membrane permeabilization, and, thereby trigger its own relase into the cytosol.
The next steps leading to apoptosis appear rather straightforward, and involve mitochondrial activation with the release of cytochrome c and activation of caspase-3, which can be prevented by overexpression of Bcl-2. Cytosolic gentamicin could act directly on mitochondria or indirectly through impairment of the proteosomal degradation of Bax (see Figure 3C).
Current studies examine the role of reactive oxygen species (ROS) formed in lysosomes by interaction of iron with gentamicin in triggering lysosomal membrane destablisation and in activating the apoptosis mitochondrial pathway.
|
These studies are now extended to other antibiotics, including polycationic peptides and saponins, as means to screen for and assess their potential to cause early toxicities.
Selected references on drug-induced apoptosis (by reverse chronological order; for full reference list, see our publication list)
Additional
information: <tulkens@facm.ucl.ac.be>
Last significant update: December 27, 2010